
The Luminescent Mating Displays of Photeros annecohenae: An Elaborate Light Show
Come nighttime, bioluminescent ostracods live up to their name.
Displays begin during the first truly dark
hour of the night when no moonlight is present,
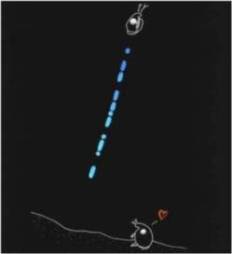 typically about
35 to 50 minutes post-sunset if there is no moon, otherwise after moonset. During this time, male Photeros annecohenae abandon their benthic
lifestyle, swim up into the water column and produce an
intricate
series of vertically-progressing flashes to attract
non-luminescent females of the same species (typically located
in the grassbed habitat). These coded trains of pulses serve to attract
the same species of females’ attention in hopes that at least one will approach
the male,
allowing for copulation
and successful mating. These displays occur two to ten
centimeters above the shallow grassbeds in which the organism
resides and produce a species-specific “train” of flashing luminescence (Torres
and
Morin 2007). The
protein luciferin and the enzyme luciferase are produced in the
light organ of the ostracod and secreted from nozzles on the
upper lip (Luminance in these organisms is always extracellular
(Morin 1986).) This causes a chemical reaction with the surrounding
water
typically about
35 to 50 minutes post-sunset if there is no moon, otherwise after moonset. During this time, male Photeros annecohenae abandon their benthic
lifestyle, swim up into the water column and produce an
intricate
series of vertically-progressing flashes to attract
non-luminescent females of the same species (typically located
in the grassbed habitat). These coded trains of pulses serve to attract
the same species of females’ attention in hopes that at least one will approach
the male,
allowing for copulation
and successful mating. These displays occur two to ten
centimeters above the shallow grassbeds in which the organism
resides and produce a species-specific “train” of flashing luminescence (Torres
and
Morin 2007). The
protein luciferin and the enzyme luciferase are produced in the
light organ of the ostracod and secreted from nozzles on the
upper lip (Luminance in these organisms is always extracellular
(Morin 1986).) This causes a chemical reaction with the surrounding
water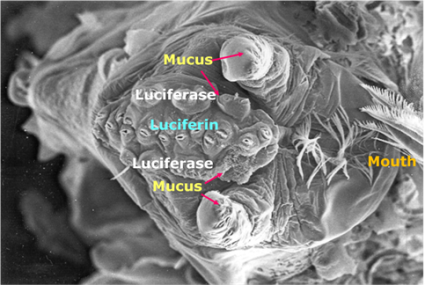 that produces flashes of bright blue light as the
male rapidly swims in the upward direction (James G. Morin 2011).
that produces flashes of bright blue light as the
male rapidly swims in the upward direction (James G. Morin 2011).
The displays
typically consist of nine to 13 pulses, three bright, longer pulses
followed by six to ten dimmer, quick pulses (Torres and Morin 2007). The length of
each train of displays is typically 50 to 70 centimeters with each
pulse lasting a second or less (Morin and Cohen 2010) and ranging from millimeters
to centimeters apart. The display tapers after 45 to 50 minutes
(Torres and Morin 2007). The train’s luminescence is typically brightest at the beginning, and, as the display progresses,
pulses become dimmer, shorter and closer together. There are two phases
of each display: an initial phase (stationary stage) and a
terminal phase (helical stage) (Morin and Cohen 2010). The
 stationary stage acts
as the “alerting phase” by attracting the attention of potential
female mates (as well as male competition). The first
signal is usually what catches a female’s attention (Rivers and
Morin 2009). This
stage produces a specific pattern of bright, spaced out flashes
and acts as a species identifier for females ready
to mate. The following helical stage is a dimmer, quicker, more
repetitive “trill” phase. It is also referred to as the
orientation phase and creates a pattern of about a dozen evenly
spaced pulses to be assessed and approached by a female (Morin
and
Cohen 2010).
Males’ displays give females an idea of their quality as
potential mates (Rivers and Morin 2009). These ostracods swim in a helical pattern
that makes them appear to be swimming more slowly and,
therefore, are more approachable to females. A receptive female
approaches a displaying, swimming male of the same species, and
the male grabs and holds her using his first pair of antennae.
stationary stage acts
as the “alerting phase” by attracting the attention of potential
female mates (as well as male competition). The first
signal is usually what catches a female’s attention (Rivers and
Morin 2009). This
stage produces a specific pattern of bright, spaced out flashes
and acts as a species identifier for females ready
to mate. The following helical stage is a dimmer, quicker, more
repetitive “trill” phase. It is also referred to as the
orientation phase and creates a pattern of about a dozen evenly
spaced pulses to be assessed and approached by a female (Morin
and
Cohen 2010).
Males’ displays give females an idea of their quality as
potential mates (Rivers and Morin 2009). These ostracods swim in a helical pattern
that makes them appear to be swimming more slowly and,
therefore, are more approachable to females. A receptive female
approaches a displaying, swimming male of the same species, and
the male grabs and holds her using his first pair of antennae.
 Other bioluminescent organisms can affect the success of
bioluminescent ostracods' mating displays by distracting females
from their own species' displays. These organisms include the
luminescent syllid polychaete worm (See photo.) and
bioluminescent dinoflagellates. These dinoflagellates also
affect displays by creating background light when a lot of them are
present in the water column (Gerrish Personal Communication).
Other bioluminescent organisms can affect the success of
bioluminescent ostracods' mating displays by distracting females
from their own species' displays. These organisms include the
luminescent syllid polychaete worm (See photo.) and
bioluminescent dinoflagellates. These dinoflagellates also
affect displays by creating background light when a lot of them are
present in the water column (Gerrish Personal Communication).
Photeros annecohenae reproduce by
copulation,
or sexual intercourse. There is variation between species in the
morphology of a male’s eighth limb (copulatory limb), and, as mentioned on
the Adaptation page, this acts as
another way to keep mating within a species. Fertilization is
internal and occurs as eggs enter the brood pouch. Two spermatophores containing sperm, produced by the male,
are placed on the female’s genetalia; a male’s
 spermatophores of
a given species are shaped to fit perfectly into the female
genetalia of that same species. It is believed that this limits the female’s ability
to mate with other males for the time being.
spermatophores of
a given species are shaped to fit perfectly into the female
genetalia of that same species. It is believed that this limits the female’s ability
to mate with other males for the time being.
Because these females are unable to mate with other
males, they retreat and
incubate their embryos inside their bivalved shells. This removes them
as potential mates, skewing the sex ratio of mating ostracods
toward males (although the sex ratio in the population as a
whole is equal (Cohen and Morin 1990)). This leads to competition between many
males for fewer receptive female mates. Females are able to
choose their mates, so males use their displays to "impress"
females. Using bioluminescence for courtship has lead to strong sexual selection in Photeros
annecohenae (Cohen and Morin 2010). This sexual selection has shaped the behavior ,
morphology and speciation of the ostracods (James G. Morin 2011). Evidence of
sexual selection includes a high sex ratio toward males, males
attracting females using luminescent signals,
sexual dimorphism
in adults and a higher parental
investment by females than males. It is believed that strong
sexual selection is ultimately what has
lead to vast diversity of species and displays in ostracods of
the Caribbean. Natural selection has lead to distinct divisions
of displays and habitats. This is useful for females in
identifying their own species' displays.
,
morphology and speciation of the ostracods (James G. Morin 2011). Evidence of
sexual selection includes a high sex ratio toward males, males
attracting females using luminescent signals,
sexual dimorphism
in adults and a higher parental
investment by females than males. It is believed that strong
sexual selection is ultimately what has
lead to vast diversity of species and displays in ostracods of
the Caribbean. Natural selection has lead to distinct divisions
of displays and habitats. This is useful for females in
identifying their own species' displays.
Sexual selection is largely due to competition between
males for
female choice of mates. The higher number of receptive males has
lead to competition between males. Competition between males has
lead to different tactics in “scoring” female mates. Males may
lead, entrain or sneak. Leading occurs when
 males initiate the
first mating display of the night. Entraining occurs when a competing male produces
a display parallel to the “leader’s” (usually about half a meter
away). This occurs frequently after the initial display and results
in many displays visible at one time. This can create a
spectacular “light show” (similar to the photo to the left) for diver spectators. Entraining may
increase mating success by attracting more females to the area
as well as increase competition between males once the females
arrive (Rivers and Morin 2009). This is similar to what is seen in
fireflies
of
Southeast Asia and North America. Lastly,
sneaking
involves a “photetically-silent” male that follows a luminescent
male’s display (spirals above him) in hopes of intercepting a
female swimming across his spiral path. Males have demonstrated
an ability to switch between these mating tactics; however, once
committed, a “sneaker” will not switch to another tactic. A
sneaker is able to switch which ostracod he is ""sneaking on" but
will never become an entrainer because he does not want to miss an
opportunity to mate. The above photo illustrates what a typical
ostracod display scene may look like. Notice evidence of
entrainment and multiple species of
bioluminescent ostracod.
males initiate the
first mating display of the night. Entraining occurs when a competing male produces
a display parallel to the “leader’s” (usually about half a meter
away). This occurs frequently after the initial display and results
in many displays visible at one time. This can create a
spectacular “light show” (similar to the photo to the left) for diver spectators. Entraining may
increase mating success by attracting more females to the area
as well as increase competition between males once the females
arrive (Rivers and Morin 2009). This is similar to what is seen in
fireflies
of
Southeast Asia and North America. Lastly,
sneaking
involves a “photetically-silent” male that follows a luminescent
male’s display (spirals above him) in hopes of intercepting a
female swimming across his spiral path. Males have demonstrated
an ability to switch between these mating tactics; however, once
committed, a “sneaker” will not switch to another tactic. A
sneaker is able to switch which ostracod he is ""sneaking on" but
will never become an entrainer because he does not want to miss an
opportunity to mate. The above photo illustrates what a typical
ostracod display scene may look like. Notice evidence of
entrainment and multiple species of
bioluminescent ostracod.
Remember that luminescent courtship displays
occur only
in the Cypridinid family of ostracods from reef systems in the
Caribbean. Females produce luminance to deter predators but do
not signal in response to males’ displays (unlike
fireflies)
(James G. Morin 2011).
These flashing displays occur nightly (and year-round) over the ostracods' grassbed habitat. Photeros annecohenae’s displays all
move upwards in a vertical direction, which is why slow water current
is necessary for successful displays. In fact, it was found that
a display occurring in a swift water current only produced three
to five pulses, while a display in no water current produced
15 to 19 pulses.
As mentioned on the Habitat page, darkness is an
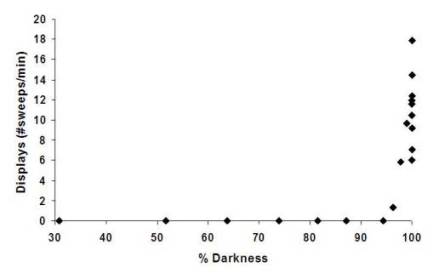
important ecological resource for Photeros annecohenae.
Reproductive (and feeding) behaviors do not begin until the “dark
threshold” is reached. This occurs when one third of the
moon or less is exposed or two to three minutes before the
beginning of nautical twilight on nights when no moon
is present (Gerrish et al. 2009). The importance of darkness can also be proven
by the fact that the number of displays increases on cloudy
nights as long as the moon is well covered and it remains dark
enough (Gerrish et al. 2009). Displays are most prevalent in the hour after sunset or
moonset. Display numbers also peak after conditions that lead to
a large number of receptive females. Females are constantly
receptive after brood release and molting; however, they mate
only when it is dark enough for mating displays, so the number
of receptive females increases during the day or during long
periods
 of too much light when mating displays cannot occur.
This is why the first hour of darkness on a night following a
full moon will have the largest number of receptive females.
of too much light when mating displays cannot occur.
This is why the first hour of darkness on a night following a
full moon will have the largest number of receptive females.
As previously-stated, bioluminescent displays are
species-specific and demonstrate a vast amount of diversity
between species. The brightness, duration of, patterns, location
and time at which they occur vary species to species.
Characteristics of individual pulses and the display train as a
whole vary as well. Most of the diversity among Photeros is a
result of differences in displays due to sexual selection and
differences of reproductive structures (Morin and Cohen 2010). There are three
overall patterns of trains of bioluminescent ostracods, but each
individual train is distinct to a given species. This allows for species
recognition, which ultimately leads to mate recognition and mate
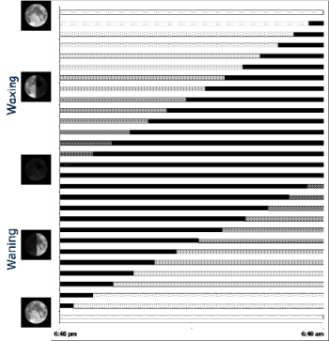
choice by females. The below sequence also shows entrainment and species-specific
displays. It illustrates how a typical series of displays would
occur.

So, a male Photeros annecohenae wowed a female Photeros annecohenae with his glowing, sexy courtship display. Now what? To learn about the life cycle of the organism, click here!