|
ABSTRACT Formation of a DNA loop by AraC proteins bound at the araI and araO2 sites, whose center-to-center distance is 211 base pairs, is necessary for repression of the araBAD promoter, PBAD, of Escherichia coli. To determine the upper and lower size limits of the loop, we constructed PBAD- reporter gene fusion plasmids with various spacings between araI and araO2 and measured their levels of expression. Spacings larger than about 500 base pairs resulted in elimination of detectable repression. No lower limit to spacing was found, suggesting that AraC protein itself possesses significant flexibility and its bending substantially aids formation of small loops. As the spacing between araI and araO2 varied, the activity of PBAD oscillated with an 11.1-base-pair periodicity implying that the in vivo helical repeat of this DNA is 11.1 base pairs per turn.
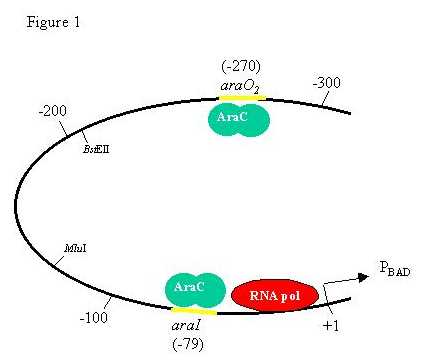
INTRODUCTION Regulation of the repressed basal level as well as the induced level of expression of the arabinose operon in Escherichia coli involves DNA looping mediated by the AraC protein, AraC proteins bound to two distinct DNA sites and bound to each other to form a DNA loop (1-4). DNA looping in the ara system requires that the two binding sites involved in forming the repression loop, araI and araO2 (Fig. 1), be on the correct face of the DNA double helix (1, 2). Although the wild-type spacing of 211 base pairs (bp) between araI and araO2 gives rise to full repression of PBAD, insertion of 5 bp between the two sites rotates one site halfway around the DNA double helix with respect to the other site and greatly interferes with repression. We interpret this finding as resulting from the torsional stiffness of DNA. |
|
Measurements on bare linear DNA made in vitro have yielded values close to 10.5 bp per turn for average DNA (6- 8). The helical repeat of DNA in vivo apparently has not been measured, but we might expect it to possess a slightly different value because on average such DNA possesses a linking number deficit. This deficit generates supercoiling (9) and, in principle, could also partially untwist the DNA. Indeed, an untwisting caused by a linking number deficit has been observed in vitro for supercoiled DNA (10, 11). Another reason for deviation from the canonical value of 10.5 bp per turn might be local supercoiling generated by transcription of adjacent genes (12). As the spacing between araI and araO2 is progressively increased or decreased, repression of PBAD should oscillate with a period about equal to the helical repeat of the DNA. Therefore, we have utilized looping in the ara system to measure the helical repeat in vivo of the DNA of the ara operon regulatory region.
Both upper and lower limits to the DNA loop size ought to exist. As the length of the DNA separating araI and araO2 increases to large values, the sites should have increasing difficulty finding one another and looping should decrease. At the other extreme, when the length of DNA separating the sites decreases, the stiffness of the DNA should hold the sites (and the proteins bound to the sites) apart and hinder loop formation. Maximal loop formation ought to occur at some intermediate loop size, much like the maximum that is seen in DNA ring closure or cyclization as a function of DNA length (5, 13, 14).
Here we report experiments showing that the helical repeat of the DNA between araI and araO2, is 11.1 bp per turn, that the upper size limit of the ara loop is about 500 bp, and that there is no lower size limit to this loop.
MATERIALS AND METHODS (abbreviated)
Construction of Insertion Plasmids. We use the distance between the apparent middle of the inverted sequence symmetries of the araI (position -59) and araO2 (position -270) sites when discussing the distance between them. Plasmid pTD3 was cut with the restriction enzyme BstEII, treated with the Klenow fragment of DNA polymerase I and alkaline phosphatase, and ligated with HaeIII fragments of E. coli chromosomal DNA smaller than 1000 bp long. The size of each insertion was determined by electrophoresis of the HindIII-EcoRl segment from the plasmids and comparison to sequenced size markers. Several candidates were chosen for further investigation by sequence analysis, isolation of repression-defective mutations, and investigation of helical twist dependency by addition of 4- or 8-bp insertions. Small-sized insertions (+13, +15, +17, +19, +21, +23, +25, +29, +31, @-33, and +35 bp) were made by inserting oligonucleotides of various sizes into the BstEII site of pLH2 and subsequent insertion of 4 nucleotides in restriction sites contained on the oligonucleotides
Construction of Deletion Plasmids. Plasmid pLH2 was cut ,with BstEII, partially digested with slow BAL-31 exonuclease (27), ligated, and transformed into strain SH322. Approximately 400 candidates were collected, and the sizes of their deletions were determined by comparing their HindIII-EcoRI fragments with known size markers. The candidates having deleted less than 200 bp, which might retain intact araI and araO2 sites, were chosen for further investigation by sequence analysis, isolation of repression-defective mutations, investigation of helical twist dependency by addition of .4-bp insertions, and in vivo footprinting.
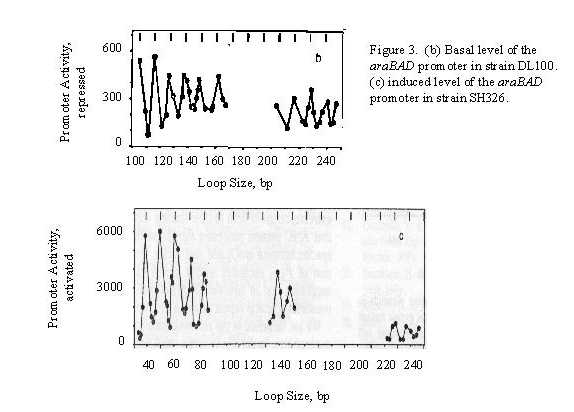
RESULTS Oscillations in repression ability as a function of spacing extended to about 400 bp loops. The hashmarks in Fig. 3b and c, which are spaced 11.1 bp apart remain in phase with the oscillations for loops 4-240 bp long, implying that the helical repeat of the DNA in vivo between the araI and araO2 is 11.1 bp per turn.
|
|