

Classification
The question of how to classify organisms has challenged biologists for centuries. Historically, specimens were grouped according to the morphological species concept—solely on their physical characteristics. Then, other ways of classification emerged. In the biological species concept, for insance, interbreeding organisms represent a species, while in the phylogenetic species concept, organisms that share a common ancestor are grouped. Additionally, the molecular species concept has been borne from our ability to do interspecies comparison via genetic analyses. Currently, we use a combination of the aforementioned concepts. Considering that there have been many ideas on how to classify life, it is not surprising that the recognized taxa (groups) we put organisms in have changed many times. Consequently, the currently recognized phylogenetic tree of all organisms is not static, and will certainly be adapted in the future as our understanding of the relationships between organisms increases. So as you read about the classification of H.peckii, bear in mind that it has changed many times in the past, and will undoubtedly change in the future.
The Domain Eukarya
The etymology of the domain Eukarya is from eu
,meaning true, and karyo, meaning nucleus; thus, all
eukaryotes have a 'true nucleus' containing their DNA. Besides a
nucleus, eukaryotes have other membrane-bound organelles such as
mitochondria and have linear DNA. In contrast, organisms from the
domains Archea and Bacteria have loose DNA in a region of the cell
called a nucleoid, lack membrane-bound organelles, and have circular
DNA (Campbell et al., 2008)..svg)
The Kingdom Fungi
The kingdom Fungi is a truly a marvelous kingdom comprised of
greater than 30,000 named species. It was once thought that fungi
were degenerate plants; however, since then, we have accumulated an
overwhelming preponderance of evidence that places fungi more
closely related to animals than plants on the evolutionary tree. The
kingdom Fungi is unique group that has many defining
characteristics, so I will name but a few.
many defining
characteristics, so I will name but a few.
First, and perhaps most significantly, all fungi have hyphae which are not like the individual cells of plants or animals you may be familiar with. Instead, hyphae are more like strands of interconnected 'cells' that are separated from each other by septa. Together, all the hyphae of an individual fungus are termed a mycelium.
Another notable distinction of fungi is the way they obtain nutrients. Fungi are heterotrophic like humans, but instead of consuming food and then digesting it, fungi excrete enzymes that digest their food externally and they consume it. To make a rough analogy, the fungal means of obtaining food would be like if humans were to secrete a special saliva to turn their their food—cheeseburgers, apples, whatever it may be—into mush, and then absorbing that slop through their skin.
Fungi also differ from animals by having ergosterol (named after the fungus ergot) instead of cholesterol in thier plasma membranes. The ergosterol in fungal plasma membranes is functionally similar to cholestrol in animal plasma membranes in that both egosterol and cholesterol provide a temperature buffer and help maintain membrane fluidity.
This is a very brief overview of what makes fungi unique. All fungi also lack vascular tissue, have cell walls made of chitin, have an alternation of generations lifecycle, and have many other defining charactistics. For further enlightenment about the kingdom Fungi, visit Tom Volk's website.
The Phylum Basidiomycota
The Basidiomycota are a diverse taxon of over 32,000 species
(Moore et al., 2011) and are commonly known as 'club fungi'
because they bear their spores externally on club-shaped structures
called basidia. In Latin, basidum means 'little pedestal'
(Campbell et al., 2008), but in the term club fungi prevailed over pedestal fungi as the common name for Basidiomycota.
The photograph below shows a magnified cross-section of gills from a
basidiomycete. The outward-facing protuberances in the photo are
basidia and the egg-shaped figures are spores.
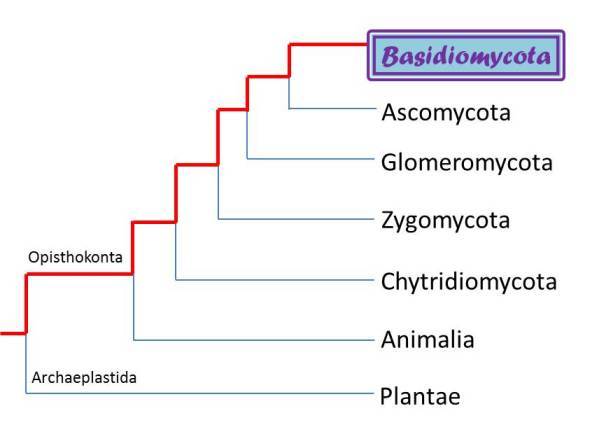
In addition to basidia, some of the Basidiomycota have clamp connections, structures unique to the Basidiomycota which allow them to maintain dikaryotic mycelia. There is a conspicuous array of morphological diversity among the fruiting bodies of Basidiomycota including gills, pores, folds, and teeth. All of these morphologies serve an identical purpose: to promote the formation and distribution of spores by maximizing surface area. Some specific examples of fungi in the phylum Basidiomycota you may be familiar with include: button mushrooms, magic mushrooms, puffballs, and polypores. The phylogentic tree below is based on DNA sequences and depicts the relatedness between five major fungal phyla as well as their association to plants and animals. As you can see, the Basidiomycota are more closely related to members of the Ascomycota such as morels and dead man's fingers than to the Glomeromycota, Zygomycota, or Chytridiomycota. In addition, the tree shows that fungi as a whole are more closely related to animals than to plants. Both animals and fungi reside within the super-group Opisthokonta and are unified by having a single posterior flagellum at some point in their development. Plants, on the other hand, belong to a separate super-group of known as Archaeplastida.
The Class Agarciomycetes
If you are a mycophagist, Agaricomycetes
would be the class of fungi
you'd choose to be stranded on a desert island with because
the vast majority of edible mushrooms reside within it. Agaricomycetes are characterized by dolipore
septa without nanopores but with perforated parenthosomes (Misra et al.,
2012). That's a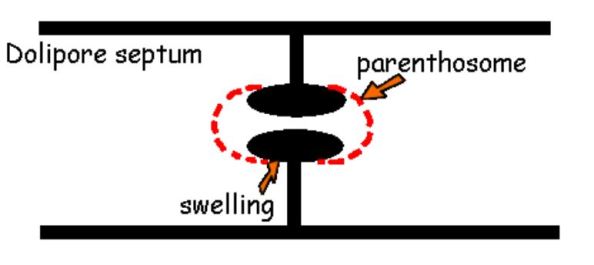 mouthful, so what does it even mean? In
the class Agaricomycetes the septa that separate individual hyphal 'cells',
have a relatively large hole in the center called a dolipore and do not have small apertures known as nanopores anywhere else in the
septa. The dolipores of Basidiomycota are enveloped in caps called parenthosomes that permit the intercellular movement of cytoplasm
while restricting the movement of large organelles (3). Although
the designation Agaricomycetes is very similar to erstwhile classifications such as Hymenomycetes and Homobasidiomycetes, recent evidence based on RNA
has enhanced our conception of fungal phylogenetics and supports the monophyly of Agaricomycetes(8).
mouthful, so what does it even mean? In
the class Agaricomycetes the septa that separate individual hyphal 'cells',
have a relatively large hole in the center called a dolipore and do not have small apertures known as nanopores anywhere else in the
septa. The dolipores of Basidiomycota are enveloped in caps called parenthosomes that permit the intercellular movement of cytoplasm
while restricting the movement of large organelles (3). Although
the designation Agaricomycetes is very similar to erstwhile classifications such as Hymenomycetes and Homobasidiomycetes, recent evidence based on RNA
has enhanced our conception of fungal phylogenetics and supports the monophyly of Agaricomycetes(8).
The Order Thelephorales
The order Thelephorales is comprised of two families: Thelphoraceae
 and Bankeraceae. Defining characteristics of the Thelephorales are ectomycorrhizae and
warted, ornamented spores.
The etymology of Theleporales comes from Greek thele
meaning 'nipple' and phor meaning 'to bear'. Add -ales,
the generic suffix for orders, and you get Thelephorales: an order
with nippled spores (Volk, T., personal communication).
and Bankeraceae. Defining characteristics of the Thelephorales are ectomycorrhizae and
warted, ornamented spores.
The etymology of Theleporales comes from Greek thele
meaning 'nipple' and phor meaning 'to bear'. Add -ales,
the generic suffix for orders, and you get Thelephorales: an order
with nippled spores (Volk, T., personal communication).
The Family Thelephoraceae

Like the order Thelephorales, the family Theleporaceae can be distinguished by warted, ornamented spores; but more specifically, the spores of Thelephoraceae are brown. On the other hand, Bankeraceae, the sister group to Thelephoraceae, have white spores. Additionally, the Thelephoraceae are exclusively tooth fungi—they all bear their spores on tooth-like projections. Magnified Hydnellum sp. teeth are pictured in the photograph to the left.
The Genus Hydnellum
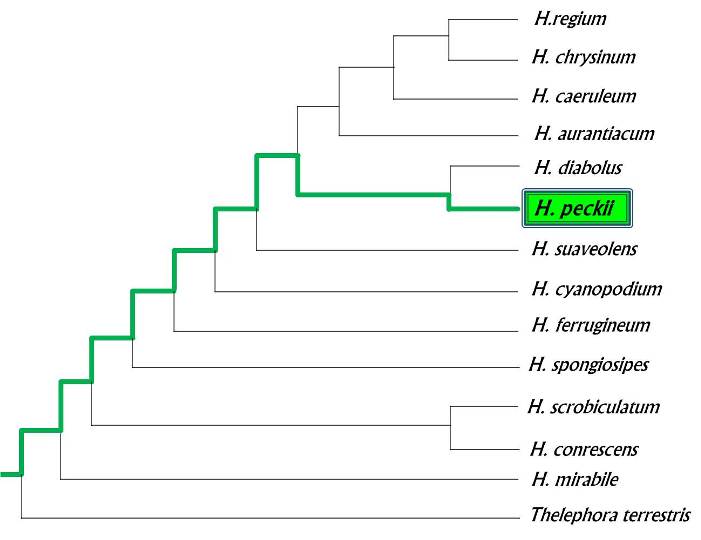
Members of the genus Hydnellum are distinguished by a "fibrous, zonate context" which means they have tough, corky flesh and coloring that may vary in zones or patches. Furthermore, Hydnellum is known for its indeterminate growth; thus, its members are able to engulf surrounding pine needles, twigs, and other matter in their path of growth. Indeterminate growth also allows individual fruiting bodies to fuse into a large fungal mass. The name Hydnellum means 'little tooth' and is in reference to the tiny 'teeth' Hydnellum bear their spores on (Westmoreland 2003). The Phylogeny below depicts relationships within the genus Hydnellum and is based on a combination of chemical sequences and morphological characteristics. H. peckii is grouped as a sister group with H. diabolus because both species share a characteristic bitter taste and have clamp connections.
The Species Hydnellum peckii
Hydnellum peckii can be identified by its white pileus (cap) that turns increasingly brown as it ages. Young, white H. peckii is considered to be tomentose, meaning it has a matted hairy texture. With age, however, H. peckii browns and its texture becomes more glabrous (smooth) and scrobiculate (pitted). Within the genus Hydnellum, morphological similarity makes it difficult to discern individual species. H. peckii has a distinct acrid, peppery taste and clamp connections— characteristics shared only by H.diabolus. Despite its stark similarity to H. diabolus, H. peckii is set apart by having a smooth, dark mature pileus in contrast to the light brown, spongy pileus of H. diabolus. Additionally, the North American distribution of H.peckii (Canada and Northern US) is more restricted than the distribution of H.diabolus which may be found along the entire East Coast from Florida to Maine. (Westmoreland, 2003).
The specific epithet peckii was chosen in honor
of acclaimed mycologist Charles Horton Peck. Peck is said to
have been a mycological autodidact who detailed more than 2,700
species of North American fungi during his lifetime. Field work was
Peck's forte. In the field, Peck illustrated and described thousands
of novel organisms. He was even know to munch on unidentified
mushroom species to determine their toxicity(6).
Now that you have learned about H. peckii's classification, I reckon you're aching learn about where H.peckii lives.