Morphology by S u n i t a N a n d i h a l l i
External Morphology

Figure 1. H. haydeniana mingling about
the leaf litter
H. haydeniana’s body is about 1.8 inches in length (Acorn & Sheldon, 2001) and has 20 twenty segments (Hopkin & Reed 1992). The surface of the body of H. haydeniana is smooth, black, and has bright yellow spots that run along the millipede’s side. This bright coloring is a warning to any predator that it is poisonous, as it secretes cyanide which is a powerful poison. (Acorn & Sheldon 2001). Although H. haydeniana is known for its chemical defenses and bright coloring, millipedes in general are mostly known for its legs. The word millipede comes from name meaning “thousand foot”. (Acorn and Sheldon 2001). The legs of millipedes grow ventrally which requires the S-shaped structure that they have, giving them the appearance of “hanging down” from their legs rather than standing on them. (Hopkin & Reed 1992). Since H. haydeniana has so many legs (two pairs per segment), they run the risk of running into each other. To combat this, H. haydeniana moves its legs in slow, coordinated, metachronal waves that start at the back of the body and move towards the head. (Acorn & Sheldon 2001). Males tend to have longer legs than females in order to more strongly grasp the females during copulation. Besides this difference between sexes, each leg is the same length throughout the body. The cuticle of each segment of millipedes consists of a dorsal tergite, ventral sternite, and lateral pleurites. In polydesmid millipedes like H. haydeniana, each segment is strengthened by the fusion of all three of these structures. (Hopkin & Reed 1992).
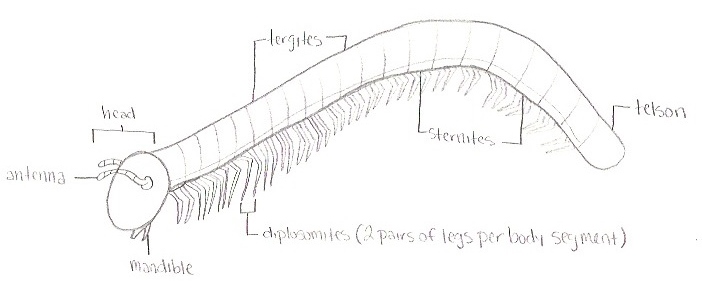
Figure 2. The head consists of organs such as the antenna and
mandibles, the body consists of tergites, sternites, and legs.
The body of H. haydeniana ends with the telson.
The cuticle
itself consists of the three layers, the very thin epicuticle,
exocuticle, and endocuticle. H. haydeniana, as well as
most other millipede species, have a calcified cuticle that they
accumulate when they eat decaying organic material. It is also
permeable to water, restricting this millipede’s habitat to
humid areas. Since H. haydeniana is an arthropod, it has
an exoskeleton and must do ecdysis. The timing of this molting
process is under hormonal control. (Hopkin & Reed 1992). On the
ventral side of body are the reproductive structures. The
gonopods of adult male millipedes are located on the seventh
segment and replace one or both legs. Female gonopods are
internal but may be extruded during copulation. When this
happens, they can be seen behind the second pair of legs. Each oviduct of female millipedes opens
separately into organs called vulvae which are in separate sacs
within the lumen and these are the structures that are everted
during copulation. (Hopkin & Reed 1992).
The telson which
is the last division of the body consist of a pre-anal segment,
a pair of anal plates, and a sub-anal scale.
The anal plates form a valve that opens during
defecation. (Blower 1985). 80 to 90 percent of dry food
ingested by H. haydeniana is excreted as feces. The two
forms of predominating nitrogenous waste in millipedes are
ammonia and uric acid. Ammonia must be excreted quickly in order
to avoid self-poisoning but uric acid can be stored temporarily
in the midgut epithelium. (Hopkin & Reed 1992).
Internal Morphology
The digestive
tract of a millipede is basically a straight tube from mouth to
anus. Small pieces of dead plant material are passed into the
lumen of the foregut where it receives secretions from salivary
glands to moisten the food. Actual digestion takes place in the
midgut, where enzymes secreted by epithelial cells break down
the plant material into its simple chemical compounds. The most
important site in any millipede for assimilation of nutrients is
the midgut epithelium. Products of digestion are absorbed by the
microvilli that border the cells in the midgut and are then
intracellularly digested and passed to the liver which is a
structure that is a layer of cells that surround the midgut.
(Hopkin & Reed 1992). Adipocytes or lipocytes, fat cells that
primarily compose of adipose tissue (specialized in storing
energy as fat), serve as part of the digestive system. They
function with the excretory system. (Camatini 1979). The organs
involved in the excretory process include the midgut epithelium,
liver, integument, exocrine glands, haemocytes, nephridial
organs, nephrocytes, ecdysial glands, malpighian tubules, and
the fat body. Millipedes have one pair of true excretory organs
called nephridial organs and regulate excretion in these
organisms. Nephrocytes are cells that take up substances in the
haemolymph and which are then metabolized. Some of these
products are then stored or returned to the circulation.
The respiratory
system in millipedes is very similar to that in insects. The
exchange of O2 and CO2 between cells in
millipedes takes place through tracheae. The tracheal system
opens up via small holes in the cuticle called spiracles (there
are two of these openings on each sternite which is
the ventral
portion of a segment of any arthropod). The openings of the
spiracles on flat-backed millipedes like H. haydeniana are
protected by a cuticular lattice (or a crystal-like structure).
Since millipedes are larger, more sluggish animals, their
respiration rates have been found to be relatively low when
compared with other arthropods. (Hopkin & Reed 1992).
Millipedes,
being arthropods, have an open circulatory system. This means
that the blood of the animal, composed of liquid and cellular
components are circulated throughout the body via the pumping
action of the heart or dorsal vessel. The liquid bathes the
organs in oxygen and nutrients and transports the products of
metabolism to and from organs of digestion, storage, and
excretion. This fluid in the body cavity is called the hemocoel.
There is no distinction between blood and interstitial fluid;
this combined fluid is called hemolymph. (Campbell et al.
2008). The principle sugar is trechalose and the main lipids are
phospholipids in the hemolymph. The blood of millipedes also
transports nitrogeneous wastes in a form that can be tolerated
and can be excreted. (Camatini 1979).
Acorn, J. and Sheldon, I. 2001. Bugs of
Washington and Oregon. Lone Pine Publishing, Edmonton, Canada.
Blower, J.G. 1985. Millipedes Keys and
Notes for the Identification of the Species. The Linnean Society
of London, London, England.
Camatini, M. 1979. Myriapod biology. Academic Press Inc., London, Great Britain.
Campbell, N.A., Reece, J.B., Urry, L.A., Cain, M.L., Wasserman, S.A., Minorsky, P.V., Jackson, R.B. 2008. Biology. Pearson Benjamin Cummings, San Francisco, California, U.S.A.